Первый критерий: что-нибудь, где-нибудь должно изменяться
Начнем с логического начала. Если память требует изменений в биохимии и структуре определенных клеток, значит, при образовании энграмм что-то и где-то в мозгу должно изменяться, но мы не знаем точно, что и где. Хуже того, анатомическое строение мозга у кур сильно отличается от его строения у млекопитающих, и до сих пор нет хорошего атласа; поэтому мне нельзя было опираться на догадки, на которые наводит знакомство с мозгом млекопитающих: у цыплят, в частности, нет того, что я мог бы назвать гиппокампом. В связи с этим для начала мне нужен был метод, который не зависел бы от локализации и механизмов изучаемых процессов. Практически любой биохимический процесс, в особенности связанный с повышением активности нейронов и синтезом в них макромолекул, требует затраты энергии. Мозг получает энергию, сжигая глюкозу; поэтому, установив, где и когда используется больше глюкозы в первые минуты после обучения, можно узнать, какая область мозга имеет отношение к хранению следов памяти. К счастью, для этого имеется довольно простой способ. Он основан на применении синтетического вещества, очень похожего на глюкозу — 2-дезоксиглюкозы (2-дГ). Если 2-дГ ввести в кровяное русло, нейроны (и все другие клетки тела) будут обмануты и начнут поглощать ее так же, как глюкозу. Внутри клетки первый же из набора ферментов, расщепляющих глюкозу, тоже примет 2-дГ за природный сахар и станет превращать ее в 2-дезоксиглюкозо-6-фосфат (2-дГ-6Ф). Это первый этап нормального расщепления глюкозы. Однако следующий по порядку фермент, который должен был бы воздействовать на глюкозо-6-фосфат, оказывается умнее и не желает иметь дело с 2-дГ-6Ф. Поэтому последний накапливается в клетках, и его количество служит мерой того, сколько они используют глюкозы. Если вводимая в кровоток 2-дГ содержит радиоактивную метку, то происходит накопление меченого 2-дГ-6Ф, и остается только измерить радиоактивность в клетке.
Согласно плану эксперимента, 2-дГ вводят цыплятам, клюющим смоченные метилантранилатом или водой бусины, выжидают полчаса, чтобы в клетках накопился 2-дГ-6Ф, забивают цыплят, извлекают и замораживают их мозг, а потом измеряют в нем радиоактивность. Но задача состоит не в том, чтобы просто выяснить, возросло ли содержание метки в мозгу обученных птенцов по сравнению с контрольными; нужно еще установить, в каком отделе мозга это произошло. Вот здесь-то и нужна методика с использованием 2-дГ. Замороженный мозг помещают в лабораторную разновидность миниатюрной мясорезки, называемую криостатом, и делают серию очень тонких срезов. Срезы переносят на предметные стекла, которые затем прижимают к листу рентгеновской пленки, заворачивают в черную светонепроницаемую бумагу и оставляют в темной комнате. После этого остается подождать несколько дней или месяцев (сроки зависят от количества радиоактивности) и проявить пленку, называемую теперь радиавтографом (а сам процесс называют радиавтографиеи).
Каждый срез оставит на пленке отпечаток, который будет тем темнее, чем больше радиоактивной метки содержалось в ткани. Степень затемненности каждого участка измеряют с помощью автоматического сканера, который своим тонким лучом прощупывает изображение и регистрирует поглощение света. С помощью компьютера черно-белое изображение можно перевести в цветное, которое легче и приятнее рассматривать, хотя цвета совершенно условны и не дают никакой дополнительной информации. Теперь можно сравнить количество метки последовательно в каждом отделе мозга контрольных и обучавшихся цыплят и попытаться выявить разницу. Я проводил такой опыт четыре сумасшедших недели в 1980 году вместе с фанатично преданной делу Маргарет Кошут — специалистом по радиоавтографии из Варшавы, а на следующий год повторил его более углубленно с нейроанатомом из Будапешта Андрашем Чиллагом, который помог идентифицировать те анатомические структуры, где Маргарет и я обнаружили изменения.
Результаты были ясны. Два участка — IMHV (Intermediate Medial Hyperstriatum Ventrale) и LPO (Lobus Parolfactorius) — «светились» у обученных животных сильнее, чем у контрольных. При этом сразу после обучения содержание радиоактивной метки было особенно высоким в левом IMHV и в левом LPO. Иначе говоря, несмотря на двустороннюю симметрию куриного мозга, состоящего, как и у млекопитающих, из двух внешне идентичных полушарий, эффект обучения был асимметричен: научением у цыплят в большей мере ведало левое полушарие [5][30].
Эти результаты были важны для нас по нескольким причинам. Во-первых, было очень интересно найти изменение в IMHV после выработки пассивного избегания, потому что Габриел Хорн раньше уже показал ключевую роль этой области мозга в импринтинге. Результаты экспериментов с пассивным избеганием и импринтингом начинали совпадать, что было хорошей новостью для обеих лабораторий. Но ни его, ни наша лаборатория не имела ни малейшего понятия, какой может быть функциональная связь (если она вообще существует) между IMHV и LPO или какова роль каждой из этих областей в деятельности мозга. Насколько мы знали, у цыплят IMHV представляет собой нечто вроде «ассоциативной коры» млекопитающих — области, где сходятся и, вероятно, интегрируются сигналы от многих сенсорных систем. Еще меньше ясности было в отношении LPO. Некоторые исследователи считали эту долю в основном «выходной» областью, координирующей двигательные реакции, в том числе клевание. По мнению других, она имела больше отношения к эмоциональным реакциям птиц, во всяком случае к чувству опасности и к ощущению неприятного вкуса (рис. 10.2).
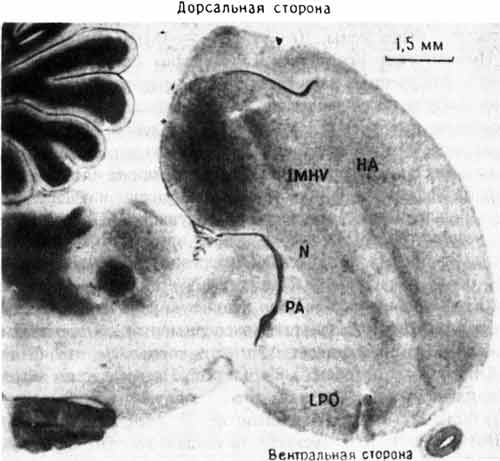
Рис. 10.2. Разрез переднего мозга цыпленка. На этой микрофотографии показано расположение IMHV и LPO. Другие отделы мозга: НА — hyperstriatum accessorium; N — neostriatum; PA — paleostriatum augmentatum. Темноокрашенная древовидная структура слева вверху — мозжечок.
Во-вторых, полученные результаты подтвердили то, о чем мы уже начинали догадываться: выраженную функциональную разницу между левой и правой частями мозга у кур. Накапливалось все больше данных о латерализации функций в мозгу у птиц; в частности, цыплята ведут себя по-разному, реагируя на предметы, которые видят правым или левым глазом [8], тогда как у певчих птиц, таких, как канарейки и зебровые амадины, «центр пения» расположен в одном из отделов левого полушария довольно близко к IMHV [9]. Но в то время мы не представляли себе, о чем могут говорить эти различия между двумя половинами мозга. Некоторые догадки появятся в конце следующей главы.
В-третьих, эти результаты имели и практическое значение: теперь мы узнали, где следует искать другие возможные изменения. Сконцентрировав внимание на IMHV и LPO и отбрасывая «ненужные» нам ткани, мы могли надеяться усилить любой изучавшийся эффект путем снижения уровня фонового шума. Обе указанные области очень малы; иссеченные из мозга, они весят не более двух миллиграммов каждая. Андраш придумал специальную пластмассовую форму, в которую мы помещали мозг, делали его срезы бритвенным лезвием, а потом тонким скальпелем вырезали под микроскопом нужные участки. Теперь мы были готовы двигаться дальше.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК