15. Биологические основы индивидуальности
15. Биологические основы индивидуальности
Благодаря экспериментам с аплизией я узнал, что изменения поведения сопровождаются изменениями силы синаптических связей между нейронами, обеспечивающими такое поведение. Но эксперименты ничего не говорили о том, как кратковременная память преобразуется в долговременную. Более того, о клеточных механизмах долговременной памяти вообще ничего не было известно.
Основанием для моих ранних экспериментов с обучением послужили аналогичные эксперименты бихевиористов, которые интересовались преимущественно механизмами приобретения знаний и их сохранения в кратковременной памяти. Долговременная память их не особенно интересовала. Интерес к ней пришел из посвященных человеческой памяти работ, предвещавших возникновение когнитивной психологии.
В 1885 году, за десять лет до того, как Эдвард Торндайк начал в Колумбийском университете свои исследования обучения у подопытных животных, немецкий философ Герман Эббингауз преобразовал изучение человеческой памяти из интроспективной дисциплины в лабораторную науку. На Эббингауза оказали влияние трое ученых: физиолог Эрнст Вебер и физики Густав Фехнер и Герман Гельмгольц, внедрившие точные методы в науку о восприятии. К примеру, Гельмгольц измерял скорость, с которой сигнал о прикосновении к поверхности кожи достигает мозга. В то время считалось, что скорость проведения сигналов по нервам неизмеримо высока и сравнима со скоростью света. Но Гельмгольц обнаружил, что она сравнительно невысока: около 90 футов[23] в секунду. Более того, время реакции, которое требуется испытуемому, чтобы отреагировать на раздражитель, оказалось еще больше, чем время, за которое сигнал достигает мозга! Это заставило Гельмгольца предположить, что значительная часть осуществляемой мозгом обработки сенсорной информации происходит бессознательно. Он назвал такую обработку «неосознанными умозаключениями» и предположил, что она основана на измерении и преобразовании нервных сигналов без осознания этих действий. Гельмгольц доказывал, что такая обработка должна обеспечиваться сигналами, определенным образом направляемыми и обрабатываемыми в различных участках нервной системы в процессе восприятия и осуществления произвольных движений.
Эббингауз, как и Гельмгольц, придерживался мнения, что психические явления имеют биологическую природу и поддаются объяснению в таких же строгих естественнонаучных терминах, что и явления физики и химии. Например, восприятие поддается эмпирическому изучению в той степени, в какой сенсорные раздражители, вызывающие ту или иную реакцию, объективны и количественно измеримы. Эббингауз решил использовать аналогичный экспериментальный подход для изучения памяти. Методы, которые он разработал для измерения памяти, по-прежнему используются.
Планируя эксперименты, посвященные механизмам запоминания новой информации, Эббингауз хотел быть уверенным, что люди, с которыми он работает, действительно приобретают новые ассоциации, а не полагаются на ассоциации, усвоенные ранее. Ему пришла в голову идея просить испытуемых запоминать бессмысленные слова, каждое из которых состояло из двух согласных, разделенных гласной (rax, paf, wux, caz и так далее). Поскольку каждое из этих слов бессмысленно, они не укладываются в уже сложившуюся систему ассоциаций. Эббингауз придумал около двух тысяч таких слов, записал каждое из них на отдельной карточке, перетасовал эти карточки и в случайном порядке доставал их, формируя списки разной длины, включавшие от семи до тридцати шести бессмысленных слов. Поставив себе непростую задачу выучить эти списки наизусть, он поехал в Париж и снял комнату в мансарде с видом на крыши прекрасного города. Там он выучил все списки по очереди, зачитывая их вслух со скоростью пятьдесят слов в минуту. Как говорила Дениз, «только в Париже человеку вообще может прийти в голову ставить такой скучный эксперимент!».
Из результатов этих экспериментов на себе Эббингауз вывел два принципа. Во-первых, он обнаружил, что памяти свойственна ступенчатость, иными словами, повторение — мать учения. Между числом повторений в первый день и объемом материала, сохранившимся в памяти на следующий, была линейная зависимость. Таким образом, долговременная память казалась просто продолжением кратковременной памяти. Во-вторых, несмотря на явное сходство механизмов кратковременной и долговременной памяти, Эббингауз отметил, что список из шести или семи слов можно выучить и запомнить с одного раза, в то время как более длинные списки требуют неоднократного повторения.
Затем он начертил кривую забывания. Он проверял сам себя через разные промежутки времени после заучивания списков, используя для каждого промежутка разные списки, и определил, какое время требовалось для того, чтобы повторно выучить каждый список с такой же степенью точности, как при первом заучивании. Он обнаружил: что-то сохраняется в памяти и после забывания, повторное заучивание старого списка требовало меньше времени и меньше повторений, чем первоначальное. И самое интересное: он обнаружил, что забывание включает по крайней мере две фазы — быстрого снижения в начале, наиболее резкого в течение первого часа после заучивания, а затем намного более пологого снижения, которое продолжается около месяца.
В 1890 году Уильям Джеймс благодаря выявленным Эббингаузом двум фазам забывания и собственной редкой интуиции пришел к выводу, что память должна включать по крайней мере два разных процесса: кратковременный, который он назвал первичной памятью, и долговременный — вторичная память. Долгосрочную он называл вторичной, потому что она предполагает вспоминание через некоторое время после первичного обучения.
Психологам, которые продолжали дело Эббиигауза и Джеймса, постепенно стало ясно, что следующий шаг в изучении долговременной памяти должен состоять в том, чтобы понять, как происходит ее закрепление (теперь этот процесс называют консолидацией). Чтобы воспоминание надолго сохранилось, необходима полная и глубокая обработка поступающей информации. Она достигается путем обращения на эту информацию внимания и осмысленной и систематической ассоциации ее со знаниями, уже хорошо закрепленными в памяти.
Первые данные, указывающие на то, что новая информация стабилизируется для долговременного хранения, получили в 1990 году два немецких психолога — Георг Мюллер и Алфонс Пильцекер. Используя методы Эббингауза, они просили группу испытуемых выучить список из бессмысленных слов достаточно хорошо, чтобы вспомнить его через двадцать четыре часа, что испытуемые из этой группы без труда и делали. Затем исследователи просили испытуемых из другой группы выучить тот же список за то же число повторений, но сразу после этого выучить еще один, дополнительный. Испытуемые из второй группы через двадцать четыре часа не могли вспомнить первый. Испытуемые из третьей группы, которым давали выучить второй список через два часа после того, как они выучивали первый, напротив, без особого труда вспоминали первый через двадцать четыре часа. Этот результат заставлял предположить, что в пределах часа после заучивания, когда первый список был занесен в кратковременную память и, возможно, в ранние стадии долговременной, память по-прежнему неустойчива. Судя по всему, для закрепления (консолидации) долговременной памяти требовался некоторый промежуток времени. После консолидации, через два или более часов, память стабилизировалась, становясь на какое-то время более устойчивой.
Представление о консолидации памяти подтверждается клиническими наблюдениями двух типов. Во-первых, с конца XIX века было известно, что черепно-мозговые травмы и сотрясения мозга могут приводить к форме потери памяти, называемой ретроградной амнезией. Боксер, который в пятом раунде получает удар по голове и переносит сотрясение мозга, обычно помнит, как он пришел на матч, но все дальнейшее стирается из его памяти. Несомненно, что ряд событий, непосредственно предшествовавших удару, записался в его памяти: волнение при выходе на ринг, движения противника во время первых четырех раундов, а возможно, даже сам удар и попытка от него уклониться, но сотрясение мозга произошло раньше, чем какие-либо из этих воспоминаний успели консолидироваться. Во-вторых, клинические наблюдения показывают, что сходная ретроградная амнезия нередко наступает после эпилептического припадка. Человек, страдающий эпилепсией, не может запомнить события, непосредственно предшествовавшие припадку, хотя припадок и не оказывает никакого воздействия на воспоминания о более ранних событиях. Это заставляет предположить, что память на ранних этапах хранения активна и легко нарушается.
Первую строгую проверку консолидации памяти провел в 1949 году американский психолог Карл Дункан. Он стимулировал электричеством головной мозг животных во время и сразу после обучения, вызывая припадок, который нарушал память, приводя к ретроградной амнезии. Когда же он вызывал у животных такой припадок через несколько часов после обучения, его действие на результаты обучения было слабым или не наблюдалось. Почти двадцать лет спустя Луис Флекснер из Пенсильванского университета сделал замечательное открытие: препараты, подавляющие синтез белков в мозгу, если принимать их во время и вскоре после обучения, нарушают долговременную память, но не сказываются на кратковременной памяти. Это открытие заставляло предположить, что для сохранения долговременной памяти требуется синтез новых белков. Обе работы, судя по всему; подтверждали представление о том, что сохранение памяти происходит по крайней мере в две стадии: кратковременная память, продолжительность которой составляет минуты, с помощью процесса консолидации, требующего синтеза новых белков, преобразуется в стабильную долговременную память, продолжительность которой составляет дни, недели и даже больше.
Вскоре были предложены новые варианты этой двухстадийной модели памяти. Согласно одной из них, кратковременная и долговременная память сохраняется в разных анатомических структурах. Но некоторые психологи, напротив, доказывали, что память хранится в одном и том же месте, просто закрепляясь со временем. Вопрос о том, требуются ли для кратковременной и долговременной памяти две отдельные структуры или они могут размещаться в одном и том же месте, принципиален для исследований поведения, особенно для исследований памяти на клеточном уровне. Было ясно, что нельзя найти ответ на этот вопрос, изучая одно лишь поведение: нужно было изучать клетки. Наши работы с аплизией давали возможность заняться вопросом, обеспечивается ли кратковременная и долговременная память одним или двумя разными нейронными процессами и происходит ли это в одном и том же или в разных участках нервной системы.
В 1971 году мы с Кэрью установили, что многократное повторение обеспечивает длительное поддержание результатов таких простейших форм обучения, как привыкание и сенсибилизация. Таким образом, эти формы обучения вполне можно было использовать для исследования различий между долговременной и кратковременной памятью. В итоге мы выяснили, что клеточные изменения, сопровождающие долговременную сенсибилизацию у аплизии, похожи на изменения, лежащие в основе долговременной памяти в мозгу млекопитающих: в обоих случаях долговременная память требует синтеза новых белков.
Нам хотелось узнать, используются ли для простых форм долговременной памяти те же места хранения (те же группы нейронов и те же наборы синапсов), что и для кратковременной. Из работ Бренды Милнер, посвященных Г. М., я знал, что у людей для сохранения сложной, эксплицитной долговременной памяти (срок хранения которой составляет дни или годы) требуется не только кора, но и гиппокамп. Но относится ли это и к более простой, имплицитной памяти? Мы с Кэрью и Кастеллуччи выяснили, что те же синаптические связи между сенсорными нейронами и мотонейронами, которые изменяются при кратковременных привыкании и сенсибилизации, изменяются и при долговременных. Кроме того, в обоих случаях синаптические изменения сопровождали наблюдаемые нами изменения поведения: при долговременном привыкании происходила депрессия синапсов, сохранявшаяся неделями, а при долговременной сенсибилизации — усиление, тоже сохранявшееся неделями. Этот результат указывал на то, что в простейших случаях кратковременная и долговременная память может храниться в одном и том же месте, в том числе для разных форм обучения.
Но оставался вопрос о механизмах. Одинаковы ли механизмы кратковременной и долговременной памяти? И если да, то какова природа процесса, обеспечивающего консолидацию долговременной памяти? Для того ям нужен синтез белков, чтобы обеспечивать долговременные синаптические изменения, связанные с долговременным хранением памяти?
Какое-то время я думал, что память может консолидироваться за счет анатомических изменений. В этом могла быть одна из причин, почему для долговременной памяти нужен новый белок. Я чувствовал, что скоро нам потребуется исследовать структуру хранения памяти. В 1973 году мне удалось взять на работу Крейга Бейли — талантливого и творческого молодого специалиста по клеточной биологии, задача которого состояла в изучении структурных изменений, сопровождающих переход кратковременной памяти в долговременную.
Бейли с его коллегой Мэри Чэнь и мы с Кэрью установили, что долговременная память не является простым продолжением кратковременной: при долговременной памяти не только синаптические изменения дольше сохраняются, но также, что более удивительно, изменяется число синапсов, действующих в нейронной цепи. А именно — при долговременном привыкании число пресинаптических связей между сенсорными нейронами и мотонейронами уменьшается, а при долговременной сенсибилизации у сенсорных нейронов вырастают новые связи, действующие, пока сохраняется память (рис. 15–1). В обоих случаях в мотонейронах происходит ряд параллельных изменений.
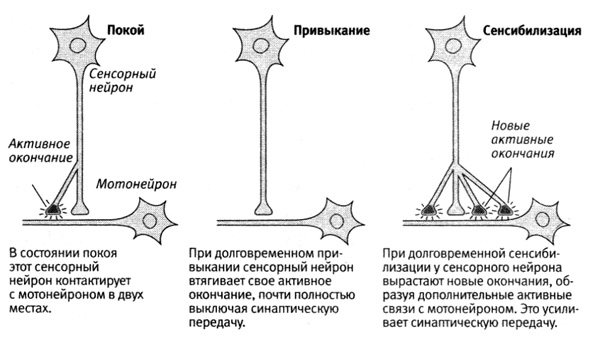
15–1. Анатомические изменения, сопровождающие долговременную память.
У этих анатомических изменений есть несколько проявлений. Бейли и Чэнь установили, что у одного сенсорного нейрона приблизительно 1300 пресинаптических окончаний, соединяющих его с примерно 25 разными клетками-мишенями — мотонейронами, возбуждающими интернейронами и тормозными интернейронами. Из этих 1300 пресинаптических окончаний лишь примерно у 40 есть активные синапсы, и только в этих синапсах имеется аппарат для выделения нейромедиатора. Остальные нейроны бездействуют. При долговременной сенсибилизации число синаптических окончаний увеличивается более чем вдвое (с 1300 до 2700), а доля активных синапсов возрастает с 40 % до 60 %. Кроме того, у мотонейрона тоже образуются дополнительные отростки, с которыми связываются некоторые новые окончания сенсорных нейронов. По мере того как память слабеет, а реакция возвращается к норме, число пресинаптических окончаний снижается с 2700 до примерно 1500, что не намного больше их первоначального количества. По-видимому, с этим остаточным явлением и связан открытый Эббингаузом факт, что при повторном выполнении того же задания животное может обучаться быстрее. При долговременном привыкании общее число пресинаптических окончаний, напротив, снижается с 1300 примерно до 850, а число активных окончаний — с 500 примерно до 100, что приводит к почти полному выключению синаптической передачи (рис. 15–1).
Таким образом, на примере аплизии мы впервые убедились, что число синапсов в нервной системе непостоянно — оно изменяется в ходе обучения! Более того, долговременная память сохраняется, пока поддерживаются связанные с ней анатомические изменения.
Эти открытия впервые позволили проверить две альтернативные теории хранения памяти. И обе оказались по-своему правильными. В соответствии с теорией одного процесса при привыкании и сенсибилизации в одном и том же месте действительно может возникать и кратковременная, и долговременная память. Более того, в обоих случаях происходят изменения синаптической силы. Но вместе с тем в соответствии с теорией двух процессов в основе этих кратковременных и долговременных изменений лежат принципиально разные механизмы. Кратковременная память связана с изменениями функций синапсов — усилением или ослаблением уже существующих связей, а долговременная требует анатомических изменений. Многократно вызываемая (повторяемая) сенсибилизация заставляет нейроны отращивать новые окончания, обеспечивая долговременную память, а многократно вызываемое привыкание заставляет нейроны втягивать уже имеющиеся у них окончания. Таким образом, вызывая глубокие структурные изменения, обучение может делать неактивные синапсы активными и наоборот.
Чтобы от памяти была польза, необходимо извлекать из нее то, что в ней записано. Извлечение информации требует соответствующих ключевых сигналов, которые животное может ассоциировать с приобретенным в ходе обучения опытом. Сигналы могут быть внешними, такими как сенсорные раздражители при привыкании, сенсибилизации и классических условных рефлексах, или внутренними — вызываемыми мыслями или побуждениями. В случае с рефлексом втягивания жабр у аплизии извлечение информации из памяти происходит благодаря внешнему сигналу, а именно прикосновению к сифону, которое и вызывает рефлекторную реакцию. Информацию об этом раздражителе извлекают из памяти те же сенсорные нейроны и мотонейроны, которые активировались при выработке этого рефлекса. Но поскольку сила и число синаптических связей между нейронами видоизменились в ходе обучения, потенциал действия, вызываемый прикосновением к сифону, достигая пресинаптического окончания, «считывает» новое состояние синапса, и извлеченная из памяти информация обеспечивает усиленную реакцию на раздражитель.
В случае долговременной памяти, как и в случае кратковременной, число изменившихся синаптических связей может оказаться достаточно большим, чтобы перенастроить нейронную цепь, но на этот раз на анатомическом уровне. К примеру, до обучения воздействие раздражителя на сенсорный нейрон аплизии может оказаться достаточно сильным, чтобы запустить потенциалы действия в мотонейронах, ведущих к жабрам, но недостаточным для запуска потенциалов действия в мотонейронах, ведущих к чернильной железе. Повторное обучение усиливает синаптические связи не только между сенсорным нейроном и мотонейронами жабр, но и между сенсорным нейроном и мотонейронами чернильной железы. После обучения воздействие раздражителя на сенсорный нейрон извлекает из памяти информацию об усиленной реакции, и это приводит к тому, что и в мотонейронах жабр, и в мотонейронах железы запускаются потенциалы действия и наряду с втягиванием жабр происходит выделение чернильной жидкости. Тем самым меняется характер поведения аплизии. Прикосновение к сифону вызывает изменение не только величины поведенческой реакции (амплитуды втягивания жабр), но и поведенческого репертуара животного.
Результаты исследований, показавшие, что нервная система аплизии физически изменяется под действием опыта, заставили нас задуматься, происходит ли то же самое и с нервной системой приматов. Изменяется ли под действием опыта человеческий мозг?
В пятидесятых годах, когда я был студентом-медиком, нас учили, что карта соматосенсорной коры, открытая Уэйдом Маршаллом, остается неизменной на протяжении всей жизни. Теперь мы знаем, что это не так. Она претерпевает постоянные изменения, определяемые опытом. Особенно подробно этот вопрос был освещен в двух исследованиях, проведенных в девяностые годы.
Первое провел Майкл Мерцених из Калифорнийского университета в Сан-Франциско. Он открыл, что у разных особей обезьян сенсорные карты коры существенно отличаются в деталях. Например, у некоторых из них кисти рук представлены в коре намного более обширными областями, чем у других. В первой работе по этому вопросу Мерцених не отделял действие опыта от генетической предрасположенности, поэтому не исключена была возможность того, что эти различия генетически предопределены.
Затем Мерцених провел дополнительные эксперименты, чтобы определить сравнительный вклад генов и опыта в эти различия. Он обучал обезьян добывать гранулы корма, прикасаясь указательным, средним и безымянным пальцами к вращающемуся диску. После нескольких месяцев тренировок области коры, связанные с этими пальцами, особенно с их кончиками, которыми обезьяна прикасалась к диску, существенно расширились (рис. 15–2). Одновременно увеличилась и тактильная чувствительность пальцев. Другие исследования показали, что тренировка различения цветов или форм тоже приводит к изменениям в анатомии мозга и улучшает навыки восприятия.
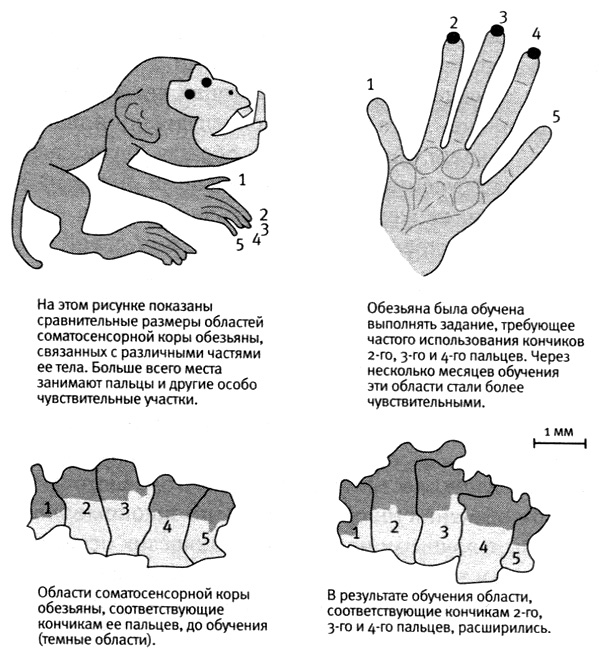
15–2. Сенсорные карты коры меняются под действием опыта. (По материалам статьи: Jenkins et al., 1990).
Второе исследование провели Томас Эльберт и его коллеги из Констанцского университета в Германии. Они сравнили томограммы головного мозга скрипачей и виолончелистов с томограммами мозга людей, не занимающихся музыкой. Музыканты, играющие на струнных инструментах, используют четыре пальца левой руки для модуляции звука струн. Пальцы правой руки, которая держит смычок, не задействованы в столь высокодифференцированных движениях. Эльберт обнаружил, что область коры, связанная с соответствующими четырьмя пальцами правой руки, у музыкантов такая же, как у немузыкантов, в то время как область, представляющая четыре пальца левой руки, в мозгу скрипачей и виолончелистов намного обширнее (более чем в два раза), чем в мозгу немузыкантов. Более того, у музыкантов, которые начали играть на скрипке или виолончели в возрасте до тринадцати лет, области коры, представляющие четыре пальца левой руки, обширнее, чем у музыкантов, которые взяли в руки инструмент после этого возраста.
Эти впечатляющие изменения коры головного мозга в результате обучения говорили о том, что закономерности, сходные с выявленными нами в анатомических исследованиях обучения у аплизии, проявляются и у других животных. Степень представленности какой-либо части тела в коре головного мозга зависит от интенсивности и сложности использования этой части тела. Кроме того, как показало исследование Эльберта, такие структурные изменения лучше проходят в ранние годы жизни. Поэтому, например, Вольфганг Амадей Моцарт стал великим музыкантом не только потому, что ему достались подходящие гены (хотя гены тоже важны), но и потому, что он начал тренировать навыки, которые принесли ему славу, в том возрасте, когда его мозг был более пластичным.
Кроме того, результаты наших экспериментов с аплизией показали, что пластичность нервной системы, то есть способность нейронов изменять силу и даже число синапсов, служит механизмом, лежащим в основе обучения и долго временной памяти. В результате, поскольку все люди растут в разных условиях и имеют разный опыт, устройство мозга каждого человека уникально. Даже у однояйцевых близнецов, у которых одинаковые гены, мозг все равно разный в связи с разным жизненным опытом. Так принцип клеточной биологии, впервые обнаруженный в ходе наших экспериментов с простым моллюском, оказался одной из глубоких биологических основ человеческой индивидуальности.
Наше открытие того, что кратковременная память возникает за счет функциональных изменений, а долговременная — за счет анатомических, поднимало новые вопросы. Какова природа консолидации памяти? Почему для этого требуется синтез новых белков? Чтобы узнать это, нам нужно было проникнуть внутрь клеток и исследовать их молекулярную структуру. Я и мои коллеги были готовы на этот шаг.
Как раз в то время мы узнали ужасную новость. Осенью 1973 года Олден Спенсер, мой лучший друг, вместе со мной основавший отделение нейробиологии Нью-Йоркского университета, начал жаловаться на слабость кистей рук, из-за которой он стал хуже играть в теннис. В течение нескольких месяцев ему поставили диагноз «боковой амиотрофический склероз» (БАС, или болезнь Шарко) — неизлечимая и всегда смертельная болезнь[24]. Узнав об этом от одного из ведущих неврологов страны, Олден впал в депрессию и стал готовить завещание, думая, что может умереть в течение недели. Но у Олдена был также артрит локтевого сустава — симптом, обычно не связанный с БАС. Поэтому я предложил ему пойти к ревматологу.
Олден пошел к очень хорошему врачу, и тот заверил его, что у него не БАС, а диффузная болезнь соединительной ткани (коллагеновая болезнь), что-то вроде красной волчанки. Когда Олден узнал этот, намного более оптимистичный, диагноз, его настроение улучшилось. Но через несколько месяцев он снова пришел к своему неврологу, и тот заверил его, что независимо оттого, есть у него артрит или нет, у него определенно БАС. Настроение Олдена сразу опять испортилось.
После этого я поговорил с его неврологом, сказал ему, что Олдену явно очень тяжело принять этот диагноз, и спросил, не может ли он помочь Олдену, подав ему какую-то надежду. Но невролог, в высшей степени достойный человек и заботливый врач, настаивал на том, что никак не может так поступить, потому что это значило бы обманывать Олдена относительно его будущего, что нечестно по отношению к нему. Он сказал: «Ведь мне нечего ему предложить. Ему просто незачем и не стоит приходить ко мне. Пусть продолжает ходить к своему ревматологу».
Я обсудил этот план с Олденом и независимо от него с его женой Дианой. Они оба решили, что это хорошая мысль. Диана была убеждена, что Олден не хочет принимать диагноз, который, как мы с ней признали, судя по всему, был верным.
В течение следующих двух с половиной лет Олдену постепенно становилось все хуже. Поначалу он передвигался с тростью, затем в инвалидном кресле. Но он все время продолжал посещать лабораторию и заниматься наукой. Несмотря на то что ему стало сложно читать лекции, он все равно преподавал, хотя и вел теперь меньше занятий. Никому из нашей группы, кроме меня, не был известен его настоящий диагноз, и никто не думал (по крайней мере, не говорил), что его болезнь — это не какая-то особая форма артрита. Олден продолжал заниматься спортом и регулярно плавал в специальном бассейне для инвалидов недалеко от дома. За день до смерти, в ноябре 1977-го, он был у себя в лаборатории и готовился участвовать в обсуждении, посвященном обработке сенсорной информации.
Смерть Олдена была сокрушительным ударом для каждого из нас, для всей нашей сплоченной группы. Мы лет двадцать общались с ним почти каждый день, поэтому теперь ритм моей жизни и работы надолго нарушился. Я по-прежнему часто думаю об Олдене.
В этом я был не одинок: мы все ценили его самоиронию, скромность, безграничное великодушие и неиссякаемые творческие способности. В 1978 году мы организовали в память о нем фонд, финансирующий курс лекций, и премию его имени, ежегодно вручаемую выдающимся ученым в возрасте до пятидесяти лет, у которых лучшие работы их жизни еще впереди. Лауреатов премии выбирает Центр нейробиологических и поведенческих исследований Колумбийского университета — сотрудники, магистранты и аспиранты, постдоки и преподаватели.
Первые годы после смерти Олдена наша работа успешно продолжалась, и со стороны можно было подумать, что все в порядке, но для меня лично это было очень тяжелое время. После Олдена в том же 1977 году умер мой отец, а в 1981-м — брат. Я принимал немало участия в заботах о них и после их смерти был не только подавлен и измотан психологически, но и обессилен физически. Большое облегчение мне всегда приносила возможность целиком сосредоточиться на работе. Непростые задачи, решения которых мы искали, и наши удивительные открытия были для меня в то время особенно ценным убежищем от горестных реалий повседневной жизни, связанных с этими невосполнимыми утратами.
Этот тяжелый период стал для меня еще горестнее в связи с отъездом сына Пола на учебу в колледж в 1979 году. Когда Полу было семь лет, я убедил его заняться шахматами и брать уроки тенниса, и впоследствии он стал неплохо играть и в то и в другое. Я томе играл в шахматы и мог поощрять его интерес к ладьям и коням. Но в теннис я не играл. Поэтому, когда мне было тридцать девять, я начал брать уроки и вскоре научился посредственно, но с удовольствием играть, что и продолжаю делать регулярно по сей день. С тех пор как Пол начал играть в теннис, он был одним из моих постоянных партнеров. К последнему классу средней школы он стал необычайно хорошим теннисистом и был моим единственным партнером. Его отъезд лишил меня не только сына, но и партнера по теннису и шахматам. Я начал чувствовать себя как Иов.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
15. Биологические основы индивидуальности
15. Биологические основы индивидуальности Обсуждение работ Гельмгольца, посвященных бессознательным умозаключениям, основано на материалах следующих публикаций: С. Frith, Disorders of cognition and existence of unconscious mental processes: An introduction, in: E. Kandel et al., Principles of Neural Science, 5th ed. (New York: McGraw-Hill,
ГЛАВА 14 Непороговые биологические эффекты
ГЛАВА 14 Непороговые биологические эффекты В годы, последовавшие за испытанием принципов «третьей идеи» в 1955 году, как нашим, так и вторым объектом были разработаны многочисленные термоядерные изделия разных весов и мощностей, предназначенные для различных носителей.
ОСНОВЫ ЭТИКЕТА
ОСНОВЫ ЭТИКЕТА Как готовиться к праздничному обеду или ужину • Занять прибывших на торжество гостей – трудная задача для хозяев. Аперитив – одно из ее решений. Чаще всего аперитив означает угощение гостей перед застольем незначительным количеством крепких десертных,
Основы
Основы Накануне Европы То было время, когда тысячи и тысячи людей, волею случая приставленных к сложным и отдаленным от врага формам борьбы, испытали внезапное желание: лечь с пулеметом за кустом, какой поплоше и помокрее, дождаться, пока станет видно в прорезь прицела —
Записки о войне основы
Записки о войне основы С. 17…. «Правда» печатала стихи от Демьяна Бедного до Ахматовой. — О Демьяне Бедном Слуцкий знал: «Жил в Кремле. Хотел — ходил к Ленину, хотел — ходил к Сталину» (см. его очерк «Н. Н. Асеев и вожди»). Об Ахматовой знал, что нелюбима советской властью и
Глава 1. Основы РОА
Глава 1. Основы РОА Нападение Германии и ее союзников 22 июня 1941 года было для Советского Союза тяжелым потрясением не только в военном, но и в политическом плане. Война разом обнажила все скрытые до сих пор внутренние противоречия советского государства. В условиях
Глава третья. Ссылка: «Мир освобожденной индивидуальности»
Глава третья. Ссылка: «Мир освобожденной индивидуальности» Кто смолоду не был социалистом, в старости будет мерзавцем. Ж. Клемансо «Один из старых северных русских городов, где все уже по-русскому: и речь русская старого уклада, и собор златоверхий белокаменный, и
Основы драматургии
Основы драматургии Познакомилась я с ним во время сессии в библиотеке. Мы сидели за одним столом. Три дня он был так увлечён своей термодинамикой, что не обращал на меня никакого внимания. Мне тоже было не до него, потому что зачёт по теории драматургии был одним из самых
Основы понимания
Основы понимания Моим первым оперативным начальником в Ахтубинске был Борис Александрович Щепанский — полненький полковник, очень хорошо известный в авиационной контрразведке Дальнего Востока и Сибири, весьма неординарная личность. Кстати, он нещадно курил и,
3.6 Теоретические основы
3.6 Теоретические основы Целый ряд «мер и мероприятий» способствовал непрерывному и неуклонному прогрессу советской ракетно–космической техники, также как и других высоких и военных технологий. Одной из таких мощнейших мер была прикладная наука. Ее воздействие на
Новые «Основы»
Новые «Основы» Возвратившись в «Узкое», Ваня заявил руководителю авторского коллектива Ф. В. Константинову, что отказывается, да и не должен писать учебное пособие, создаваемое по решению XX съезда партии, потому что получил строгое партийное взыскание. Но все:
Основы общества
Основы общества Применение Фрейдом своих открытий к ваянию, литературе и живописи было достаточно смелым, однако оно бледнеет перед его попыткой раскопать более отдаленные основы культуры. Когда мэтру было уже за пятьдесят, он поставил перед собой именно эту задачу:
КОРНИ И ОСНОВЫ
КОРНИ И ОСНОВЫ Сценическая судьба «Сказания о невидимом граде» была несчастливой. Ни в Петербурге в 1907 году, ни годом спустя в Москве опера не имела настоящего успеха. Широкая публика нашла ее вялой и чрезмерно серьезной по содержанию. Не было в ней ни очевидной для всех
Мозаика индивидуальности
Мозаика индивидуальности А если в меню попадут атомы, намагниченные другими, что будет?!Диффузия.Мозаика